| I. ABSTRACT
Human T-cell leukemia virus type
1 (HTLV-1) protease was purified from Ik19 and Nde19 pellets using affinity
chromatography. The purified protease was then refolded and activated
by dialyzing against acetate and citric buffers. Processed protease
samples with concentrations close to .08 µg/µL showed the highest
rate of enzyme activity per concentration and autoprocessing.
II. TEXT In this project, HTLV-1 protease was purified and activated using the procedures discussed below. A. Purification A Nde19 or Ik19 pellet of 1.3 ±
.3 g was first resuspended in 1 ml of buffer A (50 mM Tris, pH 7.9, 5 mM
imidazole, 300 mM NaCl) and was placed on ice to thaw. The pellet
was then sonicated using six 10 seconds pulses. The bacterial
lysate was cleared by centrifugation and 1 ml of Ni-NTA was added to the
supernatant and the mixture was incubated at room temperature with a rocker
for about two hours. The mixture was loaded on a Ni-NTA column. The column
was then rinsed with 2 ml of buffer A, washed with 20 ml of buffer B (20
mM imidazole), and eluted with 2.5 ml of buffer C (20 mM Tris, pH 7.9,
1 M imidazole, 500 mM NaCl). Small samples from each step of the
purification was collected to be loaded on a SDS-Gel later in the experiment
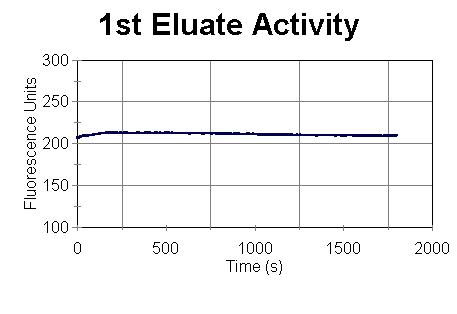
(Figure III). The eluate solution obtained at this stage has no activity.
The activity of an eluate solution was tested at this stage by using 200
µL of an eluate sample, 480 µL of buffer D (50 mM Tris, pH
7.9, 300 mM NaCl), and 5 µL of substrate. The eluate and buffer
D were first mixed in a quartz spectrophotometer cell and equilibrated
at 38 C for 10 minutes. 5 µL of substrate was then added to
the cell and was inverted to mix. The sample activity was read over
time as shown in Figure I.
Figure I: Nde 19 Prep
of 11/11/99
Since the obtained elution solution
from the column is not active, we must activate the prepared sample using
the procedures discussed below.
B. Expression and Activation To obtain active HTLV-1 protease,
the purified protein was refolded by sequential dialysis against buffer
E (10 mM sodium acetate buffer, pH 3.5, and [DTT]) and buffer F (100 mM
sodium citrate buffer, pH 5.3, 1 M NaCl, and DTT). The protease was
first dialyzed against 500 ml of buffer E for two hours and then against
a second 500 ml of buffer E for 17 hours. After the two dialysis against
buffer E, the protease was dialyzed against 500 of buffer F for 3 hours
and then against a second 500 ml of buffer F for 21 hours. Nde19
eluate solutions from pellets of approximately 1.5 g were first diluted
to about 12 ml and Ik19 eluate from pellets of the same size were diluted
to about 6 ml using buffer D before performing the dialysis against buffers
E and F. If the solutions are not diluted before the two dialysis,
large precipitates will form, and if the solutions are diluted too much,
very low concentration of protease will be obtained with low activity per
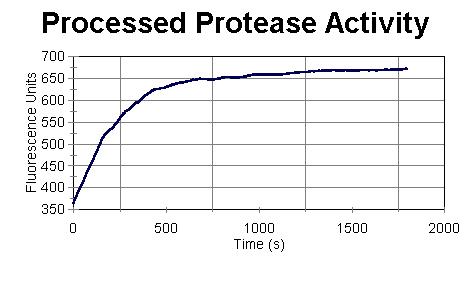
concentration. Figure II illustrates the activity of 200 µL
of the refolded Nde 19 Prep from 11/11/99. The concentration of the
sample was .0753 µg/µL.
FIGURE II: Nde 19 Prep
of 11/11/99
The refolded protease used for Figure II is highly active. The sample has cut the substrate almost completely in 1800 s. To obtain the activity per concentration, one can find the slope of the straight line before it becomes curved, and divide the slope by the concentration. The slope of the straight line obtained from Figure II is 1.064 f/s, the concentration is .07530 µg/µL, and therefore, the activity per concentration is 14.13 (f.µL)/(µg.s). The concentration of the refolded protease samples prepared using the above procedures ranged from 0.04 µg/µL to 1.2 µg/µL and the activity per concentration ranged from 5 (f.µL)/(µg.s) to 17 (f.µL)/(µg.s). The best activity per concentration was obtained when a very small amount of precipitates were formed. At that stage, the concentration of the refolded protease is around .08 µg/µL Samples from different steps of the
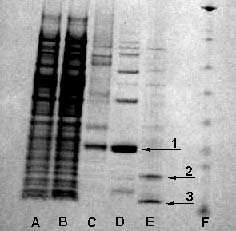
purification were analyzed by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and are shown in Figure III. A major band is
visible at 20 kDa in the elution sample (Fig. III, 1). This band corresponds
to the expected molecular size of HTLV-1 protease fused to the 27-amino-acid
pET19b his tag. The 20-kDa protein also bound to the His-Bind affinity
column and eluted with imidazole (buffer C). Therefore, we can conclude
that the band visible at 20 kDa (Fig. III, 1) corresponds to our desired
unprocessed protease.
Figure III: SDS-PAGE
analysis of samples
The N-terminal amino acid sequence
of the processed protease shows that precursor protease cuts itself at
the junction of the His tag and the first amino acid of the mature HTLV-1
protease. The amino acid sequence at this cleavage site is very different
from the protease cleavage sites seen in the HTLV-1 Gag protein and can
explain why autoprocessing is not completely efficient (Fig. III, E). (1)
III. References
1. Y. Shirley Ding, Sherry M. Owen, Renu B. Lal, and Richard A. Ikeda (1998). Efficient Expression and Rapid Purification of Human T-Cell Leukemia Virus Type 1 Protease. J Virol. 72: 3383-3386. The
World of Science. © 1997-2000, Ali Javey. All Rights Reserved.
|